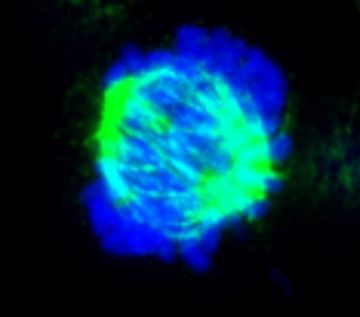
The
mitotic spindle checkpoint protects against chromosome missegregation and
aneuploidy by delaying sister chromatid separation in the presence of
unattached or improperly attached kinetochores. For this purpose, spindle
checkpoint proteins accumulate on unattached or mis-attached kinetochores to
prevent activation of the anaphase-promoting complex/cyclosome (APC/C) and
delay anaphase onset. The DNA damage response on the other hand, is a complex
network of signaling pathways that coordinates cellular reactions, ranging from
cell cycle arrest and DNA damage repair to the induction of senescence or apoptosis, to DNA lesions.
·
Studies ranging from the 1950s until recently had suggested
that the DNA damage response and the mitotic cell division are unrelated
processes with the former preserving genome stability during interphase and the
cell division machinery ensuring accurate distribution of the genetic material to
the two daughter cells. Findings from our lab were perhaps the first
to challenge this view and it is now becoming recognized that DNA
damage response proteins also play essential roles in mitosis in the absence of
damaged DNA, indicating functional interactions between these two essential mechansms that maintain genome stability.
·
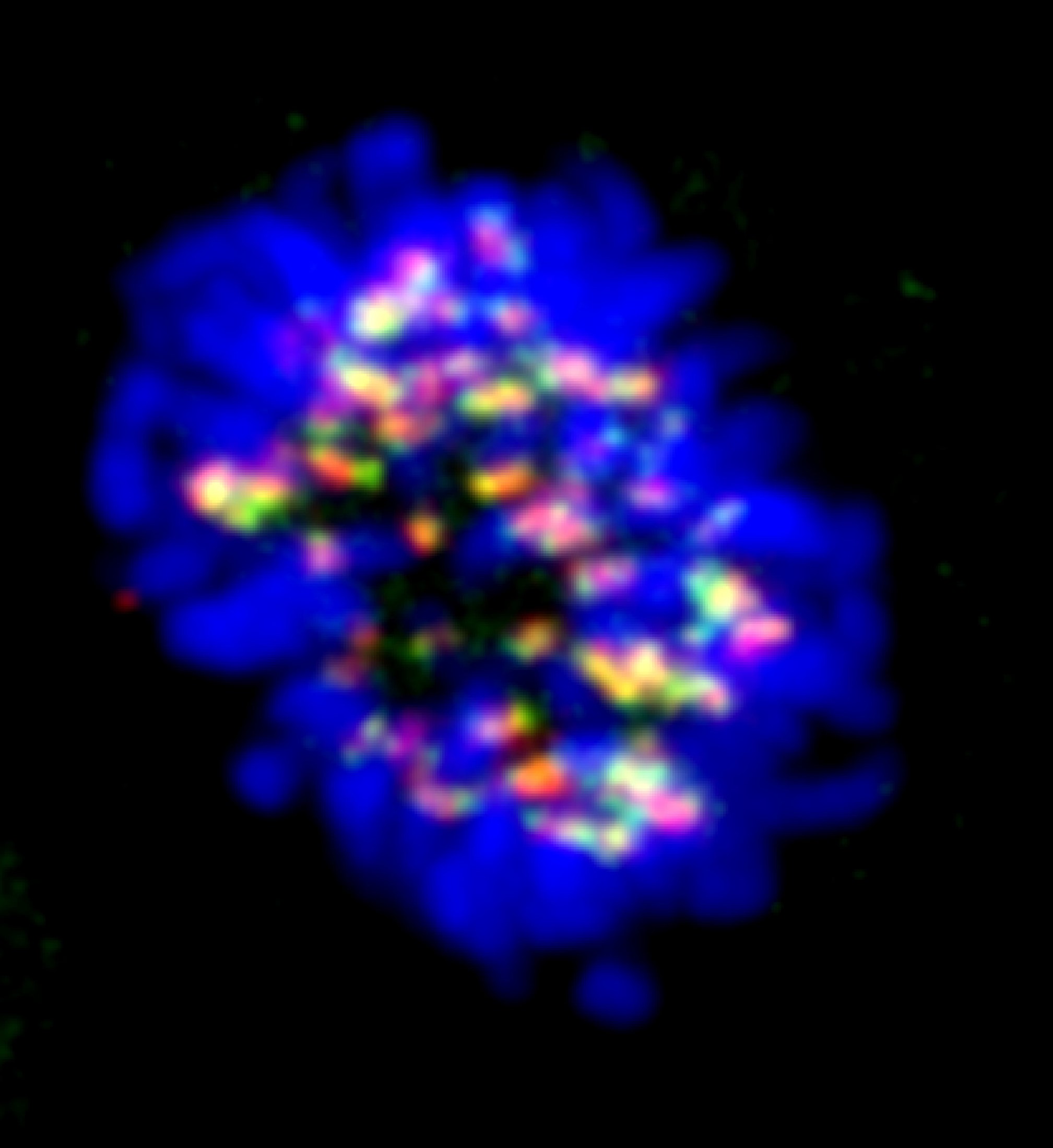
More specifically, we showed that the DNA damage response
kinases Chk1 and Chk2 promote correction of misattached kinetochore-microtubules and spindle
checkpoint signaling by phosphorylating the Aurora B kinase, an important
regulator of mitotic cell division that forms the catalytic core of the
chromosomal passenger complex (CPC), in prometaphase. We also identified serine
331 (S331) as the conserved Chk1/Chk2-site on Aurora B and determined how phosphorylation
of this site promotes Aurora B catalytic activity.
·
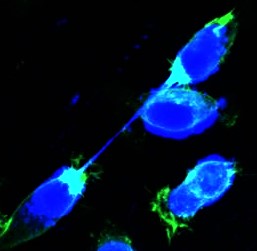
We also demonstrated that the endosomal sorting
complex required for transport (ESCRT) protein Chmp4c promotes optimal
chromosome alignment and segregation by interacting with
kinetochore-microtubules and by promoting localization of the RZZ (Rod-Zw10-Zwilch)
spindle checkpoint complex to unattached kinetochores. These findings identify unexpected connections between the membrane remodeling and mitotic machineries that remain
to be fully explored.